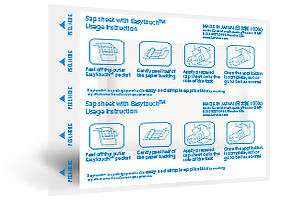




















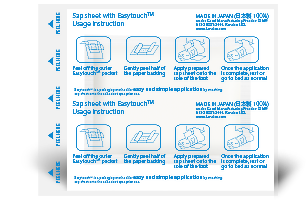
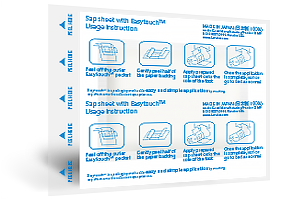
















-
 Featured Cure
Featured CureOur Dedication
Each day millions of families count on Kenrico SAP Sheet to help them be able to live the way they love.Each day millions of families count on Kenrico SAP Sheet to help them be able to live the way they love. The TRMX 4 Original is our continuous dedication to enhance the quality of life for people and their families around the world.
Learn more -
 DETOXIFY, VITALITY, INTELLIGENCE
DETOXIFY, VITALITY, INTELLIGENCEProven Results
Ensure your skin patch has SPOROLIFE to prevent weight gain and lymphedema. The inclusion of SPOROLIFE has been mandated by the European authority. Ensure your skin patch has SPOROLIFE to prevent weight gain and lymphedema. The inclusion of SPOROLIFE has been mandated by the European authority. Find out how TRMX 4 Original impregnated with Sporolife detoxifies and improves intelligence, confidence, and vitality.
Read more -
 Our Patent & Invention
Our Patent & InventionGet down to business
Kenrico shows the world why they are the pioneer and the best in their fields.Kenrico shows the world why they are the pioneer and the best in their fields. Driven by the will to save and improve life through medicines, new innovation has been achieved through Sap Sheet (PAT# 8317762), SPOROLIFE (PAT# 7182965), Lexirin (PAT# 6793945), Zeolite (PAT# 3348957).
Learn More -
 Doctors Recommended
Doctors RecommendedDiamond Hybrid Edition
Burns fat pockets and cellulite, shrinks dimpling of the skin and tightens the overlying skin with no diet, no pills, no exercising and no plastic surgery.Burns fat pockets and cellulite, shrinks fibrous bands that causes dimpling of the skin and tightens the overlying skin by stimulating collagen formation with no diet, no pills, no exercising and no plastic surgery.
Read more -

 Popular Video
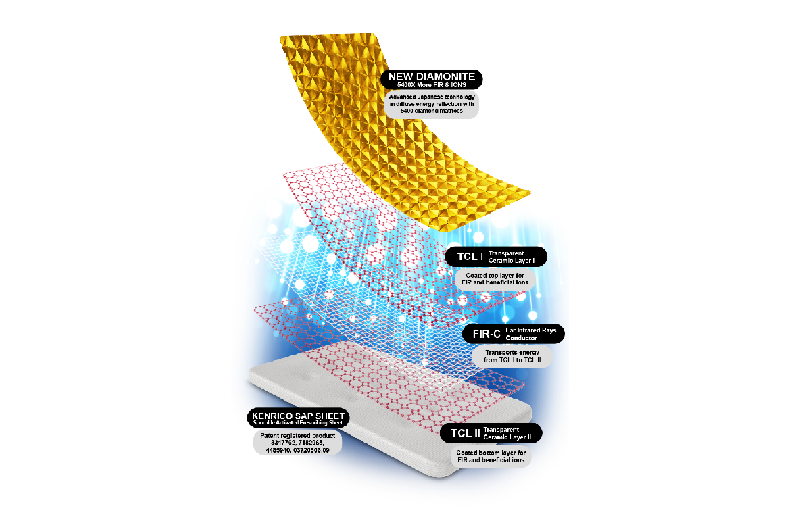
Popular VideoTRMX 5 50th Anniversary contains Sporolife and Diamonite
The safest and the most effective self-medicating pad. Clinical studies proven and doctor's recommended. The safest and the most effective self-medicating pad. Clinical studies proven and doctor's recommended.
Watch Ad -
 Our Factory
Our FactoryAhead of the curve
Kenrico factory in Japan is equipped with Japanese medical technology produced by precised Japanese workmanship.Kenrico factory in Japan is equipped with Japanese medical technology produced by precised Japanese workmanship. Kenrico has been certified with GMP, ISO 9001:2008, 14001, US FDA, Health Canada and other certifications from other countries.
Inspect our factory -
 3D Foot Reflexology Chart
3D Foot Reflexology ChartBest of both worlds
Kenrico Sap Sheet directly affects the reflex points on the feet. Get the multi-languages app free from Apple Store and Google Store now.Kenrico Sap Sheet directly affects the reflex points on the feet. Get the multi-languages app free from Apple Store and Google Store now.
Get the chart -
 Our support
Our supportKenrico Care
Medical doctors at Kenrico provides answers to your questions and prescribe treatments to help your recovery.Medical doctors at Kenrico provides answers to your questions and prescribe treatments to help your recovery. KenricoCare consists of medical doctors and pharmacists of Japanese, American and German nationals.
Get medical advice
DELAYS & PRICING FOR CT ADHESIVES FROM MAY 2024